common name: European earwig
scientific name: Forficula auricularia Linnaeus (Insecta: Dermaptera: Forficulidae)
Introduction - Distribution - Description and Life Cycle - Behavior - Hosts - Management - Selected References
Introduction (Back to Top)
The European earwig, Forficula auricularia Linnaeus, is a predatory insect with omnivorous feeding habits that occasionally result in significant injury to some economic crops. More often, they can be a nuisance by contaminating vegetables with their presence. Yet, in some cases they can be beneficial due to their predatory habits.
Figure 1. Adult male (bottom) and female (top) European earwigs, Foricula auricularia Linnaeus. The forceps help determine the sex of the adult. Photograph by Jim Kalisch, University of Nebraska-Lincoln.

Distribution (Back to Top)
The European earwig is native to Europe, western Asia, and northern Africa, but has been introduced to North America, Australia and New Zealand.
The European earwig was first observed in North America at Seattle, Washington in 1907. It spread quickly, and was reported from Oregon in 1909, British Columbia in 1919, and California in 1923. It reached Rhode Island in 1911, New York in 1912, and most other provinces and northern states in the 1930s and 1940s. Presently it occurs south to North Carolina, Arizona and southern California, but due to its preference for temperate climates it is unlikely to become abundant in the southeastern states. Also, it is not very tolerant of arid environments, but survives where irrigation is practiced. However, it is often intercepted in Florida and other southeastern states, arriving in shipments of produce and other products, as well as hitchhiking on motor homes and other vehicles (Paul Choate, personal comunication).
Other earwigs are abundant in North America, but few are as numerous as European earwig, and none are as damaging. The ringlegged earwig, Euborellia annulipes (Lucas), and the African earwig, Euborellia cincticollis (Gerstaecker), are probably the only other species of concern to crop producers. Ringlegged earwig is widespread, but African earwig is limited to southwestern states. Neither species is native to North America, though ringlegged earwig has become the dominant earwig in Florida.
Description and Life Cycle (Back to Top)
Adult: The adult normally measures 13-14 mm in length, exclusive of the pincher-like cerci (forceps), though some individuals are markedly smaller. The head measures about 2.2 mm in width. Adults, including the legs, are dark brown or reddish brown in color, though paler ventrally. The antennae have 14 segments. Despite the appearance of being wingless, adults bear long hind wings folded beneath the abbreviated forewings. Although rarely observed to fly, when ready to take flight the adults usually climb and take off from an elevated object. The hind wings are opened and closed quickly, so it is difficult to observe the wings.
Adults bear a set of cerci at the tip of the abdomen. Adults can use the cerci in defense, twisting the abdomen forward over the head or sideways to engage an enemy, often another earwig. The pronounced cerci are the most distinctive feature of earwigs; in the male the cerci are strongly curved whereas in the female they curve only slightly. The forceps of some males are 3/16 inch (5 mm) long, while others have forceps 3/8 inch (9.5 mm) long (Jacobs 2009).
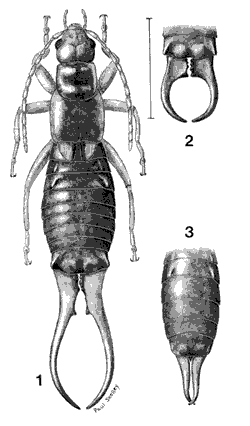
Figure 2. (1) Adult European earwig male; (2) tip of abdomen of male with short, sharply curved forceps; (3) tip of abdomen of female earwig.
Egg: The egg is pearly white in color and oval to elliptical in shape. The egg measures 1.13 mm in length and 0.85 mm in width when first deposited, but absorbs water, swells, and nearly doubles in volume before hatching. Eggs are deposited in a cell in the soil, in a single cluster, usually within 5 cm of the surface. Mean number of eggs per cluster is reported to range from 30 to 60 eggs in the first cluster. The second cluster, if produced, contains only half as many eggs. Duration of the egg stage under winter field conditions in British Columbia averages 72.8 days (range 56-85 days). The second or spring brood of eggs requires only 20 days to hatch. Eggs are attended by the female, who frequently moves the eggs around the cell, and apparently keeps mold from developing on the eggs (Buxton and Madge 1974). Females guard their eggs from other earwigs, and fight with any intruders.
Figure 3. Adult female European earwig, Foricula auricularia Linnaeusm with eggs and young. Photographed in Chester, United Kingdom. Photograph by Nabokov, en.wikipedia.org.

Nymph: The nymphal stages, four in number, have the same general form as adults except that the wings increase in size with maturity. The cerci are present in all instars, growing in size with each molt. The body color darkens, gradually changing from grayish brown to dark brown, as the nymph matures. The legs are pale throughout. The wing pads are first evident in the fourth instar. Mean head capsule width is 0.91, 1.14, 1.5, and 1.9 mm in instars 1 to 4, respectively. Mean body length is 4.2, 6.0, 9.0, and 9 to 11 mm, respectively. The number of antennal segments is 8, 10, 11, and 12 in instars 1 to 4. Mean duration (range) of instars under laboratory temperatures of 15-21°C is 12.0 (11 to 15), 10.2 (8 to 14), 11.2 (9 to 15), and 16.2 (14 to 19) days for instars 1 to 4. However, development time is considerably longer under field conditions, requiring 18 to 24, 14 to 21, 15 to 20, and about 21 days for the corresponding instars. Young nymphs are guarded by the mother earwig, who remains in or near the cell where the eggs are deposited until the nymph's second instar is attained.
Figure 4. Immature (early instar - first?) European earwigs, Foricula auricularia Linnaeus. Photograph by Cheryl Moorehead, Bugguide.net.

One generation is completed annually, and overwintering occurs in the adult stage. In British Columbia, eggs are deposited in late winter, eggs hatch in May, and nymphs attain the adult stage in August. Overwintering females may also produce an additional brood; these eggs hatch in June and also mature by the end of August (Lamb and Wellington 1975). In Washington, these events occur about one month earlier (Crumb et al. 1941). Only a single brood of eggs is produced in colder climates, such as Quebec (Tourneur and Gingras 1992).
Behavior (Back to Top)
Earwigs are nocturnal, spending the day hidden under leaf debris, in cracks and crevices, and in other dark locations. Their nighttime activity is influenced by weather. Stable temperature encourages activity, and activity is favored by higher minimum temperatures but discouraged by higher maximum temperatures. High relative humidity seems to suppress movement whereas higher wind velocities and greater cloud cover encourage earwig activity (Chant and McLeod 1952). They produce an aggregation pheromone in their feces that is attractive to both sexes and to nymphs, and release quinones as defensive chemicals from abdominal glands (Walker et al. 1993).
Social behavior is weakly developed in European earwig. Males and females mate in late summer or autumn, and then construct a subterranean tunnel (nest) in which they overwinter. The female drives the male from the nest at the time of oviposition. Eggs are manipulated frequently, apparently cleaning them to prevent growth of fungi. She will relocate the eggs in an attempt to provide optimal temperature and humidity for the eggs. Although the female normally keeps the eggs in a pile, as the time for hatching approaches she spreads the eggs in a single layer. After hatching, females continue to guard the nymphs and provide them with food. Food is provided by females carrying objects into the nest, and by regurgitation. Thus, there is parental care, but no cooperative brood care (Lamb 1976).
The most comprehensive treatment of European earwig biology is by Crumb et al. (1941), although the publications by Jones (1917) and Fulton (1924) are informative.
Hosts (Back to Top)
This insect is omnivorous, feeding on a wide variety of plant and animal matter. Although its predatory habits somewhat offset its phytophagous behavior, on occasion European earwig can inflict significant injury to vegetables, fruit, and flowers. Bean, beet, cabbage, celery, chard, cauliflower, cucumber, lettuce, pea, potato, rhubarb, and tomato are among the vegetable crops sometimes injured.
Seedlings and plants providing the earwigs with good shelter, such as the heads of cauliflower, the stem bases of chard, and the ears of corn, are particularly likely to be eaten, and also to be contaminated with fecal material. Among flowers most often injured are dahlia, carnation, pinks, sweet william, and zinnia. Ripe fruit such as apple, apricot, peach, plum, pear, and strawberry are sometimes reported to be damaged.
The European earwig is reported to consume aphids, spiders, caterpillar pupae, leaf beetle eggs, scale insects, spiders, and springtails as well as vegetable matter. Aphid consumption is especially frequent and well documented (McLeod and Chant 1952, Buxton and Madge (1976a and b, Mueller et al. 1988). In addition to the higher plants mentioned above, earwigs consume algae and fungi, and often consume vegetable and animal matter in equal proportions (Buxton and Madge 1976a).
Damage (Back to Top)
The economic status of earwigs is subject to dispute. Undoubtedly earwigs sometimes incur damage to vegetable and flower crops, both by leaf consumption and fruit injury. Foliage injury is usually in the form of numerous small holes. Tender foliage and flower petals may be completely devoured. However, the physical presence of earwigs as crop contaminants is perhaps even more important, because most people find their presence and odor repulsive. The annoyance associated with their presence is exacerbated by the tendency of earwigs to aggregate, often in association with human habitations; most people simply find them annoying.
The propensity of European earwig to consume other insects is an important element in offsetting their reputation as a crop pest. For example, they can be a significant component of the insect predator assemblage of pear and apple orchards in Washington State (Horton et al. 2002), of citrus orchards in southern Europe (Romeu-Dalmau et al. 2012), and of kiwi orchards in New Zealand (Logan et al. 2011). They have been shown to feed on various pests such as aphids, scale insects, psyllids, and midges (He et al. 2008, Logan et al. 2007, 2011). The factors limiting abundance of earwigs include insecticide use, and availability of shelters where they can avoid predation by birds (Logan et al. 2007, 2011). Use of least toxic insecticides and providing shelter for the earwigs have been suggested and evaluated. However, augmenting the earwig population by field release, and providing them with additional shelter to enhance survival, have had mixed success in suppressing pest populations (Carroll and Hoyt 1984, Carroll et al. 1985, Logan et al. 2007, He et al. 2008).
Management (Back to Top)
Sampling. Population monitoring can be accomplished with baits and traps. Small piles of baits placed among dense vegetation can be used to monitor abundance because they often attract large numbers of earwigs. Wheat bran or oatmeal can serve as a bait. Likewise, traps take advantage of the natural tendency of earwigs to hide in crevices and dark spots, and can be used to detect presence of earwigs, and to estimate abundance.
Cultural control. On residential property or in small gardens, persistent trapping can be used to reduce earwig abundance, though this approach is challenging if the initial earwig density is high. Boards placed on the soil will be attractive to earwigs seeking shelter. Even more earwigs will accumulate if there are narrow grooves or channels in the board. Moistened, rolled-up newspaper placed in the garden in the evening and disposed of in the morning makes a convenient earwig trap for home gardens. A particularly effective technique is to fill a flower pot with wood shavings and invert the pot over a short stake that has been driven into the soil. Traps can also be placed in trees because earwigs favor convenient shelters after foraging for food during the evening.
Biological control. Several natural enemies are known, including some that were imported from Europe in an attempt to limit the destructive habits of this earwig in North America. Some authors have suggested that the most important natural enemy is the European parasitoid Bigonicheta spinipennis (Meigen) (Diptera: Tachinidae), which has been reported to parasitize 10 to 50% of the earwigs in British Columbia. Others, however, report low incidence of parasitism (Lamb and Wellington 1975). Another fly, Ocytata pallipes (Fallén) (Diptera: Tachinidae) also was successfully established, but causes little mortality. Under the cool, wet conditions of Oregon, Washington, and British Columbia, the fungi Erynia forficulae and Metarhizium anisopliae also infect earwigs (Crumb et al. 1941, Ben-Ze'ev 1986). The nematode Mermis nigrescens appears to be an important mortality factor in Ontario, where 10 to 63% of earwigs were infected during a 2-year period (Wilson 1971). However, this nematode has not been reported from earwigs elsewhere. Avian predation can be significant (Lamb 1975).
Chemical control. Commercial products are rarely formulated specifically for earwigs because they rarely are a severe problem. Rather, products sold for grasshoppers, cutworms, slugs, and sowbugs are applied for earwig control. Bait is most effective if applied in the evening.
Pillbugs, sowbugs, centipedes, millipedes and earwigs
Selected References (Back to Top)
- Ben-Ze'ev IS. 1986. Notes on Entomophthorales (Zygomycotina) collected by T. Petch: II. Erynia ellisiana sp. nov., non Erynia forficulae (Giard.), comb. nov., pathogens of Forficulidae (Dermaptera). Mycotaxon 27: 263-269.
- Buxton JH, Madge DS. 1974. Artificial incubation of eggs of the common earwig, Forficula auricularia (L.). Entomologist's Monthly Magazine 110: 55-57.
- Buxton JH, Madge DS. 1976a. The food of the European earwig (Forficula auricularia L.) in hop gardens. Entomologist's Monthly Magazine 112: 231-237.
- Buxton JH, Madge DS. 1976b. The evaluation of the European earwig (Forficula auricularia) as a predator of the damson-hop aphid (Phorodon humuli). I. Feeding experiments. Entomologia Experimentalis et Applicata 19: 109-114.
- Carroll DP, Hoyt SC. 1984. Augmentation of European earwigs (Dermaptera: Forficulidae) for biological control of apple aphid (Homoptera: Aphididae) in an apple orchard. Journal of Economic Entomology 77: 738-740.
- Carroll DP, Walker JTS, Hoyt SC. 1985. European earwigs (Dermaptera: Forficulidae) fail to control apple aphids on bearing apple trees and woolly apple aphids (Homoptera: Aphididae) in apple rootstock stool beds. Journal of Economic Entomology 78: 972-974.
- Chant DA, McLeod JH. 1952. Effects of certain climatic factors on the daily abundance of the European earwig, Forficula auricularia L. (Dermaptera: Forficulidae), in Vancouver, British Columbia. Canadian Entomologist 84: 174-180.
- Choate PM. (2001). The earwigs (Dermaptera) of Florida and eastern United States. http://entomology.ifas.ufl.edu/choate/dermaptera.pdf (5 September 2019)
- Crumb SE, Eide PM, Bonn AE. 1941. The European earwig. USDA Technical Bulletin 766. 76 pp.
- Fulton BB. 1924. The European earwig. Oregon Agricultural Experiment Station Bulletin 207. 29 pp.
- He XZ, Wang Q, Xu J. 2008. European earwig as a potential biological control agent of apple leaf-curling midge. New Zealand Plant Protection 61: 343-349.
- Horton DR, Broers DA, Hinojosa T, Lewis TM, Miliczky ER, Lewis RR. 2002. Diversity and phenology of predatory arthropods overwintering in cardboard bands placed in pear and apple orchards of central Washington state. Annals of the Entomological Society of America 95: 469-480.
- Jacobs S. (January 2009). European earwigs. Penn State Entomology. (5 September 2019)
- Jones DW. 1917. The European earwig and its control. USDA Bulletin 566. 12 pp.
- Lamb RJ. 1975. Effects of dispersion, travel, and environmental heterogeneity on populations of the earwig Forficula auricularia L. Canadian Journal of Zoology 53: 1855-1867.
- Lamb RJ. 1976. Parental behavior in the Dermaptera with special reference to Forficula auricularia (Dermaptera: Forficulidae). Canadian Entomologist 108: 609-619.
- Lamb RJ, Wellington WG. 1974. Techniques for studying the behavior and ecology of the European earwig, Forficula auricularia (Dermaptera: Forficulidae). Canadian Entomologist 106: 881-888.
- Lamb RJ, Wellington WG. 1975. Life history and population characteristics of the European earwig, Forficula auricularia (Dermaptera: Forficulidae), at Vancouver, British Columbia. Canadian Entomologist 107: 819-824.
- Logan DP, Maher BJ, Connolly PG, Pettigrew MJ. 2007. Effect of cardboard shelter traps on predation of diaspidid scale insect by European earwigs, Forficula auriularia, in kiwifruit. New Zealand Plant Protection 60: 241-248.
- Logan DP, Maher BJ, Connolly PG. 2011. Increased numbers of earwigs (Forficula auriularia) in kiwifruit orchards are associated with fewer broad-spectrum sprays. New Zealand Plant Protection 64: 49-54.
- Mueller TF, Blommers LHM, Mols PJM. 1988. Earwig (Forficula auricularia) predation on the woolly apple aphid, Eriosoma lanigerum. Entomologia Experimentalis et Applicata 47:145-152.
- Romeu-Daimau C, Espadaler X, Piñol J. 2012. Abundance, interannual variation and potential pest predator role of two co-occurring earwig species in citrus canopies. Journal of Applied Entomology 136: 501-509.
- Tourneur J-C, Gingras J. 1992. Egg laying in a northeastern North American (Montreal, Quebec) population of Forficula auricularia L. (Dermaptera: Forficulidae). Canadian Entomologist 124: 1055-1061.
- Walker KA, Jones TH, Fell RD. 1993. Pheromonal basis of aggregation in European earwig, Forficula auricularia L. (Dermaptera: Forficulidae). Journal of Chemical Ecology 19: 2029-2038.
- Wilson WA. 1971. Nematode occurrence in Ontario earwigs (Nematoda: Dermaptera). Canadian Entomologist 103: 1045-1048.